
Durante el período de 1960 y los finales del siglo XX, distintos investigadores concentraron esfuerzos para comprender la regulación de la biosíntesis y el metabolismo de los Eicosanoides. Estas moléculas, de carácter autacoide, se originan de los ácidos grasos poliinsaturados de 20 átomos de C, de allí que su nombre se derive del prefijo griego EICO, veinte. Las acciones fisiológicas y en ciertas condiciones, fisiopatológicas, ejercidas por estas moléculas ocurre en un orden de concentración mmolar o menor y permanecen activas por espacios de tiempo que fluctúan entre los segundos y los minutos. En ese fructífero período del siglo XX, se logró identificar: 1- el ácido araquidónico como el principal precursor de los eicosanoides, 2- el compartimiento de los fosfolípidos de membrana como el almacén celular del sustrato precursor y 3- a las fosfolipasas, como las enzimas requeridas para liberar al ácido graso precursor, que hace posible su acceso a la maquinaria enzimática de síntesis de eicosanoides. En función de su estructura molecular surgen dos grandes grupos de eicosanoides, el que agrupa a los cíclicos o prostanoides, cuyo precursor universal es la prostaglandina H (PGH), un endoperóxido cíclico sintetizado por la enzima prostaglandina endoperóxido sintasa, mejor conocida por su acrónimo, COX, de cicloxigenasa; y el que agrupa a los lineales: leucotrienos, lipoxinas, y epóxidos entre otros, que son el producto de distintas rutas enzimáticas incluyendo a la lipoxigenas y las citocromo oxidasas. Esta revisión presenta los hallazgos más importantes en la historia de los prostanoides.
Palabras clave: Eicosanoides, prostanoides, ácidos grasos w3, isoeicosanoides.
From 1960 to the end of the XX century, efforts from different laboratories were done to understand the regulation of eicosanoids biosynthesis and metabolism. These autacoids are synthesized from 20 C polyunsaturated fatty acids, from where they got the prefix EICO, twenty. The physiologic and in certain conditions pathphysiologic actions, requires ≤ mmolar concentrations remaining active for periods of time ranging from seconds to minutes. During this productive period of the XX century it was possible to identify: 1- araquidonic acid as the main eicosanoids precursor, 2- the membrane phospholipids compartment as the cellular storage of the precursor fatty acid and 3- the phospholipases as the enzymes required to liberate the precursor fatty acid rending possible its access to the eicosanoids biosynthetic machinery. Two main groups of eicosanoids arrive on the scene, classified in function to its molecular structure, one that includes the cyclic or prostanoids, where the prostaglandin H (PGH) is the universal precursor, PGH is a cyclic endoperoxide synthesized by the enzyme prostaglandin endoperóxido synthase, better known by its acronym COX for cycloxygenase, and the other, linear eicosanoids:: leukotrienes, lypoxines, and epoxides which are products synthesized from different enzymatic pathways including the lypoxygenase and cytochrome oxidases. The most relevant findings in the history of the prostanoids are considered in this review.
Key words: Eicosanoids, prostanoids, w3 fatty acid, isoeicosanoids.
Los prostanoides, conforman un grupo de autacoides (del griego autos: propio y akos: alivio, sustancia formada metabólicamente por un grupo de células que altera la función de otras células a nivel local). Se encuentran ampliamente distribuidos en el organismo y prácticamente todas nuestras células son capaces de sintetizarlos, pero la misma, se restringe a la forma de tejido-célula-específica. La expresión de la maquinaria enzimática para la síntesis de un prostanoide en particular puede restringirse a un tipo celular en un tejido específico, así en la célula endotelial (CE) de la macrovasculatura la prostaciclina I2 (PGI2), es el prostanoide mayoritario; mientras que en la plaqueta lo es el tromboxano A2 (TXA2). No obstante, estas moléculas también pueden sintetizarse de forma transcelular, un mecanismo de "cooperatividad" celula-célula, mediante el cual las células de un determinado microambiente, incorporan sustratos o metabolitos sintetizados por sus vecinas y viceversa. La síntesis de TXA2 a partir de PGH2 sintetizada por la célula endotelial es el ejemplo clásico para ilustrar el metabolismo transcelular (1-4).
El carácter autacoide de estas moléculas, reposa en su cualidad de mensajero intercelular; esta propiedad originalmente se describió como "Gewebshormone" término alemán que se uso para definir la capacidad de transferir información, entre dos o más tipos celulares, en un órgano. La síntesis y la difusión de los autacoides al medio intersticial les permite acceder a los receptores expresados por células vecinas (acción paracrina), estimular sus propios receptores de membrana (acción autocrina) o desde el compartimiento intracelular acceder a los receptores nucleares (acción intracrina). Los eicosanoides no se almacenan y tienen una vida media muy corta, 30s para el TXA2 y 2 min. para la PGI2; características como estas, los hacen estratégicos en la regulación del microambiente celular y la homeostasis. Sin embargo, en condiciones fisiopatológicas como en la hipersensibilidad inmediata, por ejemplo, la sobreproducción puede amplificar en segundos el daño celular. Esto último, generó el interés terapéutico de inhibir su síntesis, no obstante, la inhibición puede al mismo tiempo provocar la pérdida de la homeostasis en otros sistemas, conduciendo a una disfunción y a la posibilidad de daño tisular (5-8).
Los prostanoides tienen una historia muy particular, el bloqueo farmacológico de su producción, al usar aspirina como medida terapéutica, antecedió en más de 70 años a su descubrimiento. El mecanismo de acción de la aspirina o ácido acetilsalicílico, analgésico-antiinflamatorio y antipirético, utilizado empíricamente en Egipto y en la Grecia antigua; y de manera formal desde 1899, permaneció oscuro hasta que se logró identificar el mecanismo biosintético de los prostanoides. Muy posterior al descubrimiento y aislamiento de las prostaglandinas por Euler von US en 1934 (9), Vane JR logra definir, en 1971, que el bloqueo de la síntesis de los prostanoides es lo que explica las acciones farmacológicas del ácido acetilsalicílico (10). Ulteriormente, se definen los mecanismos moleculares del bloqueo mediado por la aspirina, lo que consiste en la acetilación irreversible de la serina-530 ubicada cerca del sitio activo de la prostaglandina endoperóxido sintasa (PGHS), esta enzima es la responsable de la síntesis del precursor de todos los prostanoides, la Prostaglandina H (PGH). La acetilación, mediada por la aspirina, impide que el sustrato entre al sitio activo de la enzima, en consecuencia, se bloquea la biosíntesis de los prostanoides (11). Sin embargo, las vías biosintéticas de otros eicosanoides, mediadas por las enzimas lipoxigenasas y citocromo oxidasas, quedan por el contrario favorecidas, de forma secundaria al bloqueo por aspirina (12).
El término prostanoide incluye a las prostaglandinas, las prostaciclinas y los tromboxanos.
Los prostanoides pertenecen al grupo de los eicosanoides, derivados hidroxilados de los ácidos grasos poliinsaturados (AGPI) de 20 átomos de C; los AGPI precursores se restringen a la familia de los ácidos grasos esenciales (AGE) w6 y w3. Los eicosanoides son sintetizados in-vivo por distintas rutas metabólicas, algunos compuestos son formados por más de un mecanismo enzimático. Las vías principales incluyen las mediadas por la:
Una vía No-enzimática genera productos isoméricos a los eicosanoides, los isoeicosanoides. A diferencia de los eicosanoides, estas moléculas se generan, in situ, en los Fosfolípidos (FL) de membrana por el ataque de radicales libres. La peroxidación, mediada por los radicales libres, ocurre sobre el ácido graso (AG) en la posición sn-2 del FL desde donde se libera el isoeicosanoide sintetizado, escindido por la acción de una fosfolipasa A2.
El interés biomédico por los isoeicosanoides y en particular por los isoprostanos, moléculas isoméricas a los prostanoides, reposa en su cualidad biomarcadora de la peroxidación lipídica, de allí su potencial uso como un indicador bioquímico del grado de daño celular inducido por los radicales libres. Una variedad de moléculas, los isoprostanos de las series PGD2, PGE2 y PGF2a, resultan de este proceso de peroxidación no-enzimática. Esta clasificación, surgió en función de su semejanza con las estructuras de las prostaglandinas (PGs), D2, E2 y F2a, respectivamente. Los miembros de la serie PGF2a son los más comunes, entre ellos, el 8epi-PGF2a representa la especie mayoritaria producida durante el daño oxidativo (13). En individuos aparentemente sanos, fumadores o no, el 8-epi-PGF2a se encuentra aumentado en los fumadores (14) asimismo se describe un aumento en la excreción urinaria de este isoprostano en pacientes homocigotos para la hipercolesterolemia familiar (15). Esta molécula, es capaz de producir una fuerte vasoconstricción vascular, renal y pulmonar así como de estimular la mitogénesis de las células musculares lisas de la vasculatura (VSMC) (16), este metabolito del ácido araquidónico (AA) puede igualmente sintetizarse en mínimas cantidades, por la acción de la PGHS (17) pero esta vía no es relevante como indicador de daño oxidativo.
La concentración plasmática de los isoprostanos aumenta en patologías como la aterosclerosis, la Diabetes Mellitus o el Alzheimer, desórdenes asociados al stress oxidativo (18-20).
El descubrimiento de los prostanoides y su identificación estructural comienza en los albores del siglo XX. El primer hallazgo se debe a Kurzrok R y Lieb CC en 1930 (21) médicos gineco-obstetras quienes informan que durante la ejecución de las inseminaciones artificiales, el útero, en algunos casos sufría una contracción violenta y en otros una relajación; describen con la ayuda de Sara Ratner, quien no aparece como autor, un compuesto en el líquido seminal de bajo PM capaz de producir contracción uterina. En la misma década, y de forma independiente Goldblat MW (22) señala que el semen humano contiene un factor que reduce la presión arterial y estimula al músculo liso, al mismo tiempo von Euler US, en 1934 describe que el contacto de la esperma humana fresca con el útero humano provocaba tanto una fuerte contracción como la relajación del tejido. Posteriormente usando esperma de ovejo, logra demostrar que el factor activo de estimulación del músculo liso se encontraba en la fracción de los ácidos grasos del extracto lipídico de la esperma; denominó al factor, Prostaglandina, pensando que su origen era exclusivo de la próstata (9,23,24). De inmediato sus propiedades fisiológicas y farmacológicas a muy bajas concentraciones fueron sugeridas; sin embargo, hubo que esperar cerca de 25 años para que el aislamiento, la purificación e identificación de estas moléculas fuese técnicamente posible.
Para 1962 se contaba con la estructura molecular de seis de las prostaglandinas: PGE1, PGF1a, PGE2, PGF2a, PGE3, PGF3a, y además con la certeza de que su producción era posible por otros tejidos (25-31). Los investigadores observaron que estas moléculas de 20 átomos de C contenían dobles enlaces en configuración cis, localizados en posición similar a la de algunos AG Esenciales (AGE); lo que condujo a pensar que éstos podrían ser los precursores. La hipótesis fue, simultáneamente demostrada, por estos investigadores del Karolinska Institute en Suecia y por otro grupo del Unilevers Research Laboratories de Holanda. Usaron ácido dihomo-g-linolénico (20:3w6) isotópicamente marcado, y constataron la producción de las PGE1 y PGF1a marcadas. Enseguida, siguieron los experimentos con los AGE, AA (20:4 w6) y eicosapentaenoico (20:5 w3) pudiéndose demostrar que éstos, son en efecto los precursores de las prostaglandinas de la serie 2 y 3, respectivamente (32-34). Con estas evidencias, fue fácil pensar que debía existir un endoperóxido intermediario, lo que demostraron, en 1973, al aislar los endoperóxidos PGG2 y PGH2, y al evidenciar la capacidad de estas moléculas de inducir de forma rápida e irreversible la agregación plaquetaria (35).
Sobre la base de estos hallazgos, siguieron una serie de experiencias donde se incubó una suspensión de plaquetas previamente lavadas, con AA marcado. En estas condiciones, se aislaron tres metabolitos mayoritarios: (1) el ácido 12L-hidroxi-5,8,10,14-eicosatetraenoico (l2-HETE), (2) el ácido 12L-hidroxi-5,8,10-heptadecatrienoico (HHT) y (3) un derivado hemiacetal, el ácido 8(1-hidroxi-3-oxopropil)-9-12L-dihidroxi-5,l0-heptadecadienoico, es decir, el Tromboxano B2, que posteriormente se definió como el metabolito estable del Tromboxano A2 (36,37) En función de lo anterior, quedó establecido que los prostanoides son en efecto, ácidos grasos hidroxilados (AGH) de 20 átomos de C, arreglados en una estructura cíclica de dos cadenas laterales, que contiene, una: la extremidad carboxi-terminal (COOH) y la otra, la extremidad metilénica (CH3) donde ocurre el grupo hidroxilo sobre el C 15. Por consiguiente, los prostaoides pueden ser considerados como AGH cíclicos. De acuerdo a la configuración de la parte cíclica los prostanoides pueden dividirse en:
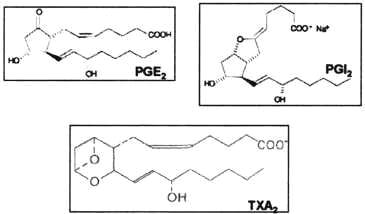
Los prostanoides están presentes en todos los tejidos animales donde se sintetizan a partir de los siguientes AG:
La enzima PGHS, puede actuar sobre cualquiera de estos sustratos generando la prostaglandina H (PGH), cuya serie será función del sustrato utilizado (Figura 2). La PGH dará origen a:
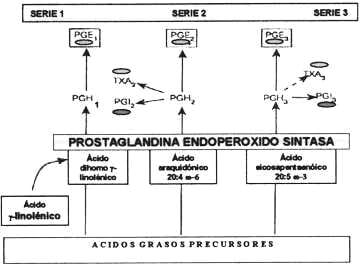
Los ácidos grasos de 20 C w6 u w3 son sustratos de la Prostaglandina Endoperóxido Sintasa (PGHS) cuyo producto la prostaglandina H (PGH) es el precursor universal de los prostanoides. La PGHS, genera, en función del AG precursor, una, de las tres series de los prostanoides. El tipo de prostanoide sintetizado dependerá del repertorio enzimático del compartimiento celular implicado. (-------->)
Molécula de menor bioactividad. La letra indica el tipo de prostanoide; la principal diferencia estructural entre estas moléculas es el número de dobles enlaces, lo que se indica por el número en subíndice, como 2 (dos dobles enlaces) para la PGE2 que caracteriza a la serie.
Los prostanoides participan en diversas funciones que incluyen la coagulación sanguínea, la ovulación, la iniciación del trabajo de parto, el metabolismo óseo, el desarrollo y crecimiento del sistema nervioso, la citoprotección del tracto gastro-intestinal; la secreción de mucus, fluidos y bicarbonato duodenal, la reparación de tejidos, la función renal, el tono vascular y la respuesta inmune. Estas acciones le otorgan un potencial farmacológico importante a estas moléculas como lo demuestra el uso de análogos de la prostaciclina, potentes antihipertensivos usados con éxito en la hipertensión pulmonar (38,39). Por el contrario, la inhibidión farmacológica de la síntesis de PGs por antiinflamatorios no esteroideos, es un factor etiopatogénico de úlcera gástrica, efecto secundario más común, consecuencia de la disminución de la síntesis gástrica de PGI2, PGE2 y PGE1 y así de la secreción de mucus, bicarbonato y del flujo sanguíneo local. Las PGE1 y E2 inhiben igualmente, la secreción ácida gástrica producida por diferentes estímulos (gastrina, histamina); lo que en suma, reduce la citoprotección y favorece el daño de la mucosa gástrica (40).
Los prostanoides pueden tener efectos antagónicos, la PGE2 provoca la relajación del músculo liso bronquial así como la dilatación de los vasos sanguíneos, mientras que la PGF2a tiene efectos contrarios en los mismos tejidos. Las prostaciclinas (PGIs) sintetizadas por las CE son potentes inhibidores de la agregación plaquetaria y producen vasodilatación lo que disminuye la presión arterial. El TXA2, aislado por primera vez de las plaquetas sanguíneas, estimula la activación y la agregación plaquetaria, uno de los mecanismos esenciales en la reparación de los tejidos. Además, este prostanoide induce la contracción de la pared arterial elevando la presión arterial. El fino balance entre las acciones antagónicas de estos mediadores es esencial para la homeóstasis vascular. Recientemente se ha considerado que la sobreproducción de TXA2 en el tejido placentario pudiera estar implicada en el mecanismo etiopatogénico de la pre-eclampsia (41). Asimismo, los resultados de un estudio de correlación en una población japonesa han permitido proponer que un polimorfismo repetido en la región del promotor del gen de la prostaciclina sintasa (PGIS) es un factor de riesgo para el desarrollo de hipertensión esencial (42).
Mecanismos de señalizaciónLas acciones biológicas de los prostanoides son mediadas por receptores de membrana heptahelicoidales acoplados a proteinas G, codificados por genes distintos (43) Los receptores (EP2, EP4), IP y DP para Prostaglandina E, Prostaciclinas, y Prostaglandina D respectivamente, median un aumento en el AMPc, a este grupo de receptores se les ha denominado "relajantes" mientras que los receptores TP (para Tromboxano A2), FP (para Prostaglandina F), y EP1 (para Prostaglandina E) se les ha agrupado como receptores "contráctiles" dada su capacidad de promover la movilización del Ca2+. Un cuarto tipo de receptor para la Prostaglandina E, el EP3, induce una disminución del AMPc por lo que se le ha denominado "inhibitorio".
Además de estos efectos, mediados por receptores de superficie en la membrana celular, se proponen acciones nucleares para algunos prostanoides como la PGJ2, un metabolito de la PGD2 y la PGI2, al funcionar como agonistas endógenos de los Receptores Activadores de la Proliferación de Peroxisomas-PPARs- (44-46). Los PPARs son una familia de factores de transcripción que regulan, entre otros, la homeostasis de los lípidos y de la glucosa (47). La activación de estos receptores por sus ligandos conduce a la heterodimerización con el receptor retinoide X (RXR) lo cual induce a su vez, la transcripción génica. Muy recientemente, Feige y col., (2005) (48) demuestran que los PPARs y los RXR pueden heterodimerizar eficientemente en ausencia de ligandos.
Posterior al aislamiento de los endoperóxidos de prostaglandinas y al descubrimiento del TXA2, Moncada en 1975 se interesa por encontrar fuentes distintas a la plaquetaria, capaces de producir TXA2. Los resultados fueron desalentadores; pero una observación importante ocurrió. Al incubar la pared de un vaso arterial con el endoperóxido precursor del TXA2, no se detectaba el producto esperado; sin embargo, el precursor desaparecía sugiriéndole a este investigador, bien la inactivación del endoperóxido precursor o bien la formación de una nueva sustancia activa. Con estos experimentos, Moncada descubría una nueva vía metabólica que conducía a la producción de una sustancia inestable capaz de provocar la relajación de las tiras de las arterias mesentéricas y celíacas, usadas en los experimentos. Rápidamente pensó, que sí en estos tejidos, en lugar de encontrar TXA2, agregante plaquetario potente y vasoconstrictor, encontraba una sustancia más bien vasodilatadora, la ecuación se completaría si esta nueva molécula era capaz de inhibir la agregación plaquetaria, lo que enseguida se demostró. A esta sustancia la denominó inicialmente PGX. (49). Posteriormente, se determinó su estructura química, la cual corresponde a un derivado éter enólico. La PGX compuesto antiagregante y vasodilatador se renombró Prostaciclina (50).
Las CEs sintetizan PGI2 a partir del AA y la secretan a la sangre y al espacio sub-endotelial alcanzando la capa media de la pared vascular. Su fijación a la plaqueta por la vía de un receptor específico, IP, activa a la enzima adenilato ciclasa, lo que conduce a la formación de adenosin monofosfato cíclico (AMPc). El AMPc inhibe el cambio de forma, la secreción, la agregación plaquetaria así como la unión del fibrinógeno a su receptor de membrana. La PGI2 tiene entonces un poder anticoagulante. La vida media de la PGI2 es relativamente corta, alrededor de 2 min., que puede prolongarse hasta 30 min. al unirse a la apolipoproteína A-1. La síntesis y la secreción de PGI2 son moduladas por un gran número de sustancias (trombina, histamina, ADP, ATP, citocinas, entre otras), por las variaciones en la fuerza mecánica del flujo sanguíneo (shear stress) así como por la alteración de la integridad del endotelio.
Las CEs pueden convertir, gracias al metabolismo transcelular, a la PGH2 de origen plaquetario, por ejemplo, en PGI2. El endotelio produce una cantidad basal de PGI2, que ha sido estimada en función de la excreción urinaria de su metabolito estable, la 2,3-dinor-6-oxo-prostaglandina F.
La aspirina, la ciclosporina y los anti-inflamatorios no-esteroideos inhiben la síntesis de PGI2 al bloquear la síntesis del precursor PGH2 por la inactivación de la PGHS.
Las células inmuno-competentes son productoras activas de prostaglandinas (PGs) y al mismo tiempo responden, de forma autocrina, al autacoide secretado. Las PGs modulan la intensidad y la duración de las respuestas inflamatoria e inmune (51-54). Entre las PGs, la PGE2 ha sido una de las más estudiadas. La PGE2 produce un número de efectos que pueden aumentar o mitigar una condición pro-inflamatoria. La PGE2 en función del tipo celular estimulado, puede: inducir fiebre, aumentar la permeabilidad vascular y la vasodilatación, favorecer la aparición de edema y la algesia mediada por los péptidos y aminas vasoactivas como la bradikinina y la histamina.
Todo ello, amplifica la condición pro-inflamatoria. Al mismo tiempo, la PGE2 inhibe la producción de citocinas pro-inflamatorias claves como el TNF, la IL- 1 e IL-6 (55,56); regulando la producción de citocinas sintetizadas por el sistema monocítico-macrofágico y por los linfocitos. Es bien conocida la inhibición sobre la proliferación de células T que ejerce la PGE2 in vitro (57) y la acción directa o indirecta sobre el patrón de producción de linfocinas, [interferon-g (IFN- g) e interleucina-2 (IL-2)] fundamentales para generar una respuesta del tipo Th1 (celular), o las IL-4, -5, e IL-10 marco de un patrón de respuesta Th2 (humoral) (58-60).
La variedad de receptores para la PGE, (EP1, EP2, EP3 y EP4) puede activar o disminuir, en función del receptor implicado, la síntesis de segundos mensajeros claves en la traducción de señales. La vía PGE2-EP4 es capaz de estimular, in-vivo, la respuesta a antígenos epidermales al favorecer la movilización, migración y maduración de las células Langerhans epidermales. Esto vincula de forma específica a la estimulación EP4 con la activación de las células Langerhans que emigran y maduran al expresar las moléculas CD80 y CD86, coestimuladoras en el proceso de presentación antigénica a células CD45RA+ (61).
El tromboxano A2, descrito inicialmente en 1975 por el grupo de Samuelsson en el Karolinska Institute, ejerce sus acciones al activar al receptor TP. La principal ruta de señalización mediada por el receptor TP es la asociada a la activación de la Fosfolipasa C (FLC) por la vía de una proteína Gaq con la subsiguiente movilización de Ca2+ y activación de la PKC (62-66). En el humano, se expresan dos receptores para el TXA2, el TPa clonado inicialmente en tejido placentario y en la línea celular, parecida a plaquetas, MEG-01 (67) y el TPb clonado a partir de una librería de cDNA de CE de la vena umbilical humana (68-69). Ambos son el producto de un gen único localizado, en el humano, en el cromosoma 19p13.3 (68).
Las isoformas a y b son idénticas excepto por el dominio citoplasmático C-terminal donde el TPa presenta un sitio de fosforilación para la proteina cinasa A (PKA), la Serina 329 y posee sólo 15 vs 79 residuos de a.a presentes en el receptor TPb. Esta diferencia entre los dominio C-terminales, esta asociada al mecanismo de "apagado" o desensibilización del receptor y a la regulación de la síntesis de AMPc. En este sentido, Hirata y col., en 1996 (64) sobreexpresando en células embrionarias de riñón humano (HEK) las isoformas a y/o b del receptor TP, demuestran que la producción de AMPc se afecta de forma opuesta. La isoforma TPa se asocia a una Gs lo que conduce al aumento de AMPc mientras que el receptor TPb se acopla a una Gi lo que inhibe a la adenilato ciclasa. (64), al mismo tiempo, el TPa y no el TPb se asocia a una Gh y por tanto a la vía PLC-fosfatidilinositol específica. No obstante, ambas isoformas, se acoplan indistintamente a las proteínas Gq y G12 (70).
En cuanto al mecanismo de desensibilización del receptor TP, las isoformas difieren en su respuesta a los prostanoides inhibitorios, PGI2 y PGD2. La señalización mediada por estas moléculas vía el receptor IP y DP, respectivamente conduce a la activación de la PKA. La fosforilación del residuo de Ser-329 blanco de la PKA resulta en la desensibilización del receptor TPa,. La isoforma TPb contrariamente no es sensible a la inactivación mediadas por PGI2 o PGD2 (71).Un nuevo orden de aspectos fisiológicos y fisiopatológicos relacionadas con el receptor TP surgen con estos hallazgos. El receptor TP humano se expresa en distintos tipos celulares incluyendo las VSMC y las CE, estas células expresan la isoformas, a y b del receptor TP; mientras que, en la plaqueta prevalece la expresión del receptor TPa.
En el circuito vascular, el TXA2 es un potente estimulador de la agregación plaquetaria y causa vasoconstricción, mientras que la PGI2 inhibe la agregación plaquetaria y causa vasodilatación. El TXA2 representa fundamentalmente un factor activador y la PGI2 el antagonista correspondiente que permite el regreso al estado de reposo no-activado, las acciones de estos prostanoides se restringen primariamente al microambiente intercelular. Como lo indican estos hallazgos, el hecho de que el receptor TPa y no el TPb se inactive por un mecanismo dependiente de la PGI2 sugiere fuertemente que la isoforma TPa es fundamental en la hemostasia vascular.
El receptor TP, puede también activarse por un ligando distinto al TXA2 como el isoprostano, 8-epi-PGF2a Este metabolito del AA, parecido a la prostaglandina F, resulta como ya mencionamos, de la peroxidación del AG mediado por radicales libres. En condiciones de stress oxidativo, el 8-epi-PGF2 puede alcanzar el orden mmolar requerido para la activación de ambas isoformas (a y b) del receptor TP, lo que ha sido demostrado in vitro (72,73). Al igual que en la biosíntesis de los prostanoides, el AG precursor de los isoprostanos no se restringe al AA, para estos compuestos en función del AGPI precursor se han propuesto nuevas clasificaciones (18,74).
La dieta es un factor que define en buena parte la composición de AG de los fosfolípidos de las membranas celulares. La composición de la dieta occidental tanto de los aceites como de los productos derivados de la carne, favorece a la familia de AG w6. El AA es el sustrato principal para la síntesis de los prostanoides y da origen a los prostanoides de la serie-2. Sin embargo, cuando se ingieren cantidades importantes de AGPI de la familia w3, como ocurre en las poblaciones que habitan en las zonas costeras, consumidoras de pescados y sus aceites, se favorece la síntesis de prostanoides de la serie-3.
Como se muestra en la Figura 2, ambas familias de AGE w3 y w6 pueden dar origen a productos similares. Es de hacer notar, que existe competencia entre estos AG, como sustratos de la reacción de la PGHS. Así, la cantidad de una familia particular de AGPI en el fosfolípido de membrana definirá el aumento relativo de una determinada serie de prostanoides. En el caso de un régimen dietario abundante en pescados y productos marinos, la elevación será a expensas de la serie-3. En general, la actividad biológica de los prostanoides de la serie-3 es menor que la de la serie-2, lo cual tiene interés terapéutico, en particular en el caso del TXA3 que carece de capacidad agregante plaquetaria mas no así la PGI3, esta prostaciclina conserva sus acciones biológicas (75). Al mismo tiempo, estos AGPI w3 inhiben la expresión de la PGHS (76) y la capacidad adhesiva del endotelio (77) lo que puede ser relevante en ciertas condiciones patológicas como los procesos inmuno inflamatorios.
El trabajo científico producido durante los últimos setenta años, permitió comprender la relevancia bioquímica, nutricional, fisiológica, fisiopatológica y farmacológica de los AGE como precursores de los eicosanoides. Estas moléculas, han explicado muchos de los mecanismos fisiopatológicos responsables de las alteraciones observadas en los trabajos pioneros de Burr y Burr (78,79) al usar dietas libres de grasas.
El estudio de los eicosanoides ha permitido evidenciar la importancia de los microambientes en un órgano o tejido y los finos mecanismos de comunicación intercelular. La especificidad de la síntesis de estos autacoides en un microambiente determinado, genera a expensas del mismo sustrato, una gama de acciones biológicas en función del tipo celular implicado.
En este sentido, el AA puede dar origen a prostanoides con acciones antagónicas, como la PGI2 y el TXA2 sintetizados por las CE y las plaquetas, respectivamente. Por otra parte, la competitividad entre los miembros de la familia de AGE, w6 y w3, por la maquinaria enzimática de los prostanoides, permite la posibilidad de síntesis de las series 2 y 3 de los prostanoides, por lo tanto, un cambio en el patrón bioquímico que favorezca a la serie 3 puede incidir positivamente en la salud pública, al limitarse la capacidad protrombótica (relación TXA2/TXA3 disminuida) mientras que, la capacidad antitrombótca y vasodilatadora se mantiene (PGI2, PG I3). Las implicaciones de la relación dietaria de AGE w6/w3 en la respuesta autacoide en un microambiente de iológica de los eicosanoides y en particular, los prostanoides.