
La tilapia (O. mossambicus x O. urolepis hormorum x O. niloticus x O. aureus) cultivada en Venezuela representa un recurso importante debido a su bajo costo y alto contenido proteico. Sin embargo a fin de garantizar su aprovechamiento racional, este producto debe llegar hasta el consumidor en óptimas condiciones de frescura. Este requerimiento ha estimulado la investigación en el efecto del retardo en la refrigeración sobre la estabilidad de la tilapia durante el almacenamiento. El objetivo de este estudio fue evaluar el impacto de tres tiempos de retardo en la refrigeración, TRR, (0 h, 4 h, 8 h) sobre la estabilidad de la tilapia almacenada en hielo (00C ± 30C) durante 21 días, mediante determinaciones de pH, Nitrógeno Básico Volátil Total (NBV-T), concentraciones individuales de nucleótidos (adenosín monofosfato, AMP; inosina monofosfato, IMP; inosina, INO; hipoxantina, Hx). También se evaluaron los cambios sensoriales durante 21 días de almacenamiento refrigerado por medio de una escala descriptiva. Los resultados mostraron que independiente del TRR considerado, se produjo una rápida caída de pH al comienzo del período de almacenamiento siendo ésta mas rápida en el pescado almacenado luego de un TRR de 8 h, en comparación con aquél almacenado con TRR de 4 h y 0 h respectivamente. El NBV-T alcanzó valores cercanos a 28 mg N% al final del almacenamiento, para cada condición de TRR considerada. El IMP se degradó progresivamente y mas rápidamente (P<0,05) a medida que aumentó el TRR, ocurriendo simultáneamente una acumulación de Hx junto con pequeñas cantidades de INO. Los atributos sensoriales dependieron del TRR evaluado (P<0,05); las tilapias almacenadas inmediatamente después del sacrificio (TRR= 0 h) tuvieron una vida útil de 21 días, en comparación con aquéllas con un TRR de 4 h y 8 h cuya vida útil fue de 15 y 12 días, respectivamente. Para cada condición de TRR la tilapia parece acumular hipoxantina (Hx) paralelamente con la degradación de inosina monofosfato (IMP).
Palabras clave: Tilapia, frescura, estabilidad en hielo, productos de degradación del ATP
The tilapia (O. mossambicus x O. urolepis hormorum x O. niloticus x O. aureus) farmed in Venezuela represents an important resource due to its low price and high protein content. However in order to guarantee its rational use, this product should reach the consumer under optimal conditions of freshness. This requirement has stimulated research on the effect of refrigeration delay on the stability of tilapia during storage. The objective of this study was to evaluate the impact of three refrigeration delay times, RDTs, (0 h, 4 h, 8 h) on the stability of the tilapia stored in ice (00C ± 30C) for 21 days, by means of pH determinations, Total Volatile Basic Nitrogen (TVBN), concentrations of individual nucleotides (adenosine monophosphate, AMP; inosine monophosphate, IMP, inosine, INO; hipoxanthine, Hx). A descriptive scale also evaluated sensory changes during 21 days of refrigerated storage. Results showed that independently of the RDT considered, a fast PH drop (P< 0.05) took place at the beginning of the storage period, being more rapid in fish stored after a RDT of 8 h, as compared to fish stored after RDTs of 4 h and 0 h respectively. The TVBN at the end of the storage period reached values near to 28 mg N% for each condition of RDT. The IMP degraded progressively and faster (P< 0,05) as the RDT increased, along with an accumulation of Hx together with small quantities of INO. The sensory attributes depended on the RDTs evaluated (P <0,05); the tilapias stored immediately after the sacrifice (RDT = 0 h) had a shelf life of 21 days, in comparison with those with RDTs of 4 and 8 h whose shelf lives were of 15 and 12 days, respectively. The tilapia seems to accumulate hipoxanthine (Hx) parallely to the degradation of the inosine monophosphate (IMP), for each RDT considered.
Key words: Tilapia, freshness, stability in ice, ATP degradation products.
El sector productivo nacional ha orientado la tecnología hacia el descubrimiento de rubros acuícolas económicamente rentables, basándose en las excelentes condiciones agro ecológicas, características de nuestro país, para la explotación de diversas especies. En este sentido, las tilapias han sido los peces exóticos de mayor éxito en la piscicultura mundial, específicamente en los países del tercer mundo por ser una especie tolerante a una cadena de condiciones ecológicas, además de tener gran aceptación a nivel del consumidor.1
Los cambios postmortem que ocurren luego de la evisceración del pescado pueden causar pérdida de los atributos sensoriales, incremento de los procesos autolíticos, proliferación bacteriana, desarrollo de rancidez, y cambios físicos que consecuentemente reducen la calidad del producto. Los métodos químicos han sido propuestos como indicadores de frescura y contaminación, sin embargo la evaluación sensorial es el método usado rutinariamente para evaluar la aceptabilidad del pescado.2
Existen numerosos estudios en el ámbito nacional sobre la estabilidad del pescado durante el almacenamiento en hielo 3-5 sin embargo hay poca información pertinente al patrón de cambios postmortem de la tilapia y su estabilidad durante el transporte y comercialización. El presente estudio tuvo como objetivo evaluar la frescura y la vida de almacenamiento en hielo de la tilapia sometida a diferentes tiempos de retardo en la refrigeración (TRR), simulando las condiciones comerciales a las que son sometidos los ejemplares durante el transporte y el almacenamiento.
Ejemplares de tilapia (Oreochromis spp) con un peso corporal promedio de 244,59 g + 27,03 g y una longitud promedio de 19,72 + 0,74 cm, obtenidos de una finca piscícola ubicada en Boca de Aroa, Edo. Falcón, fueron capturados con chinchorro y sacrificados mediante incisión cerebral.
Se evaluaron dos lotes de aproximadamente 80 individuos cada uno. Cada lote se dividió en tres grupos los cuales fueron sometidos a los siguientes tiempos de retardo en la refrigeración (TRR): 0 h, 4 h y 8 h, a temperatura ambiente (270C ± 30C), luego de lo cual cada grupo fue almacenado en cavas a temperaturas reguladas de 00C + 3 0C (hielo) y transportadas al laboratorio de productos pesqueros en el Instituto de Ciencia y Tecnología de Alimentos de la Universidad Central de Venezuela. Se tomaron muestras del músculo dorsal de tres ejemplares seleccionados al azar, para cada tiempo de retardo en el almacenamiento, cada tres días para hacer determinaciones físicas y químicas y diariamente para el análisis sensorial, por un período de tres semanas.
a) Determinación de humedad, grasa cruda, cenizas totales, proteína cruda, pH: según métodos A.O.A.C. 6 b) Determinación de nitrógeno básico volátil total (NBV-T): según el método de Pearson. 7 c) Determinación de compuestos de degradación del ATP (AMP, IMP, INO, Hx): según método propuesto por Iwamoto et al., 8con las siguientes modificaciones: 5 g de músculo dorsal homogeneizados en frío con 15 ml de ácido perclórico al 10 %, en un homogeneizador "Sorvall" Omni-mixer17.105. El homogenato obtenido fue centrifugado a 4.000 r.p.m. Por 10 min. En una centrífuga sorvall rcb-2 dotada de un rotor ss-34. Se separó el sobrenadante y se ajustó a un PH entre 6.0-7.0 con hidróxido de potasio al 50%. El precipitado formado por este procedimiento se separó centrifugando a 4.000 r.p.m. Durante 10 min. Por último, se tomó el sobrenadante y se aforó a 50 ml con agua destilada para determinar la concentración del AMP, IMP, INO e HX. La concentración de nucleótidos, se hizo sobre una alícuota de 15 µl, inyectada en un cromatógrafo líquido de alta resolución (HPLC) marca "Waters" compuesto por una bomba modelo 510, un detector UV-VIS (190-486 nm.) ajustado a 254 nm y 0.5 AUFS.
La separación de las diferentes fracciones de los nucleótidos se realizó con una columna novapack c-18 (4 mm x 3.9 mm x 15 cm) y se empleó un sistema isocrático de solventes en el que se utilizó una fase móvil compuesta por buffer fosfato PH 4,2-4,3, a una velocidad de flujo constante de 0.4 ml/min. Como patrón se preparó un estándar múltiple a partir de AMP, IMP, INO, Hx de alta pureza (sigma chem., co), con concentraciones de 20 µg/ml para AMP, IMP e INO respectivamente, y 8 µg/ml para Hx y, un volumen de inyección de 15 µl. Para el cálculo de las concentraciones de los nucleótidos se procedió al análisis de las áreas de los cromatogramas de las muestras respecto a las áreas del patrón.
Se realizó tanto en las muestras crudas como en las cocidas, empleando una escala descriptiva graduada de 0 a 10 puntos según Huss,9 indicando 10 puntos completa frescura, 8 puntos buena calidad, 6 puntos indica que el pescado es neutro en sus propiedades organolépticas y 4 puntos es el nivel de rechazo, para cada condición de tiempo de retardo. El examen del pescado crudo comprendió la evaluación de la apariencia general, una descripción de la piel, branquias, vísceras, integridad muscular, espina dorsal, ojos y órganos internos. Para la evaluación de las muestras sometidas a cocción se realizó una prueba descriptiva del olor, sabor, color y textura. La cocción se realizó colocando los filetes en bolsas de polietileno durante un período de 15 min a la temperatura de ebullición del agua.
Se realizó un análisis de varianza multifactorial (ANOVA) con un nivel de significancia de 95%, así como pruebas de corre- lación lineal, utilizando el programa STATGRAPHICS 6.0.10
Análisis proximal
El Cuadro 1 muestra la composición proximal porcentual de los dos lotes muestreados. Valores promedios de 78,43% de humedad, 17,43% de proteína, 1,03% de grasa y 1,26% de ceniza, caracterizaron el músculo dorsal de los ejemplares de tilapia empleados en el estudio. No se encontraron diferencias significativas (p> 0,05) en las variaciones de los componentes entre los dos lotes.
Cambios de pH
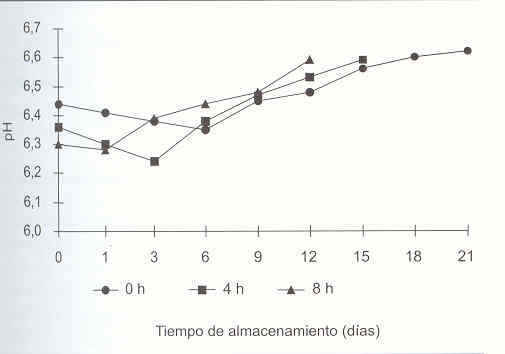
En la Figura 1, se observa que durante el inicio del almacenamiento e independiente de la condición de retardo considerada (0 h, 4 h, 8 h) se presentó una primera etapa caracterizada por la disminución de pH que fue mas acelerada en las tilapias almacenadas con un mayor TRR. El valor mínimo de pH se alcanzó en seis días en las tilapias almacenadas en hielo inmediatamente después de la captura, 6,35 en comparación con los ejemplares almacenados con TRR de 4 h y 8 h que alcanzaron valores mínimos de PH después de tres días 6,24 y un día de almacenamiento,6,28 respectivamente.
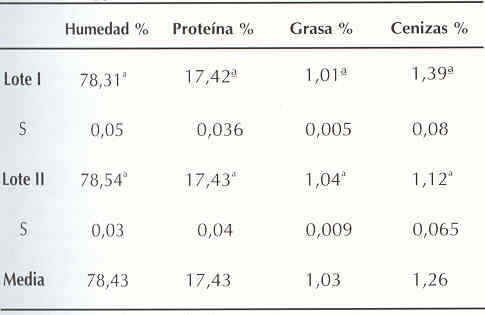
Los resultados son el promedio de 3 ó 4 determinaciones. Letras distintas en una misma columna indican diferencias estadísticamente significativas (P < 0,05).
En una segunda etapa se observó un incremento de pH en las tres condiciones de TRR evaluadas, ocurriendo éste mas tempranamente a mayor TRR: el día 3 en los ejemplares almacenados con un TRR de 8 h en comparación con el día 6 y el día 9 para los ejemplares con TRR de 4 h y 0 h respectivamente. Los incrementos en los valores de pH coincidieron, en todas las condiciones de retardo, con un aumento en la concentración del nbv-t así como con la pérdida de aceptabilidad organoléptica (pH vs nbv-t, r0 h= 0,98; r4 h = 0,98; r8 h= 0,96).
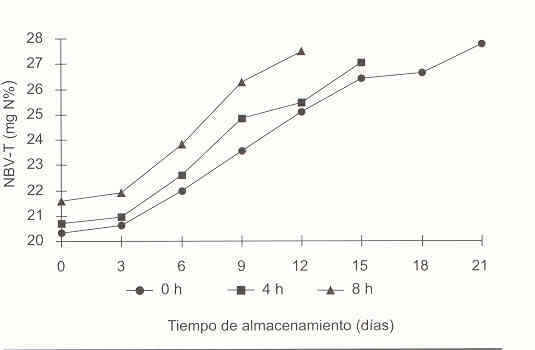
En la Figura 2 se observa que el NBV-T incrementó al avanzar el almacenamiento y al aumentar el TRR (p< 0,05). Al final del período de almacenamiento los valores de NBV-T fueron cercanos a 28 mg n%, en las tres condiciones de retardo consideradas, no alcanzando el límite permitido en productos pesqueros de 30 mg n%.11
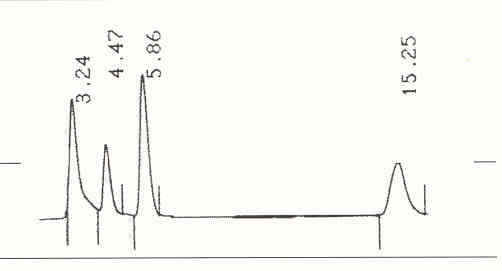
La Figura 3 muestra el cromatograma de una mezcla de los nucleótidos AMP, IMP, Hx e INO separados. El IMP es el primer analito en eluir de la columna a los 3,24 min seguido de la Hx a los 4,47 min, el AMP a los 5,86 min y la INO a los 15,25 min.
Las variaciones en las concentraciones de AMP, IMP, INO e Hx tuvieron un comportamiento similar en los tres TRR considerados (Figura 4).
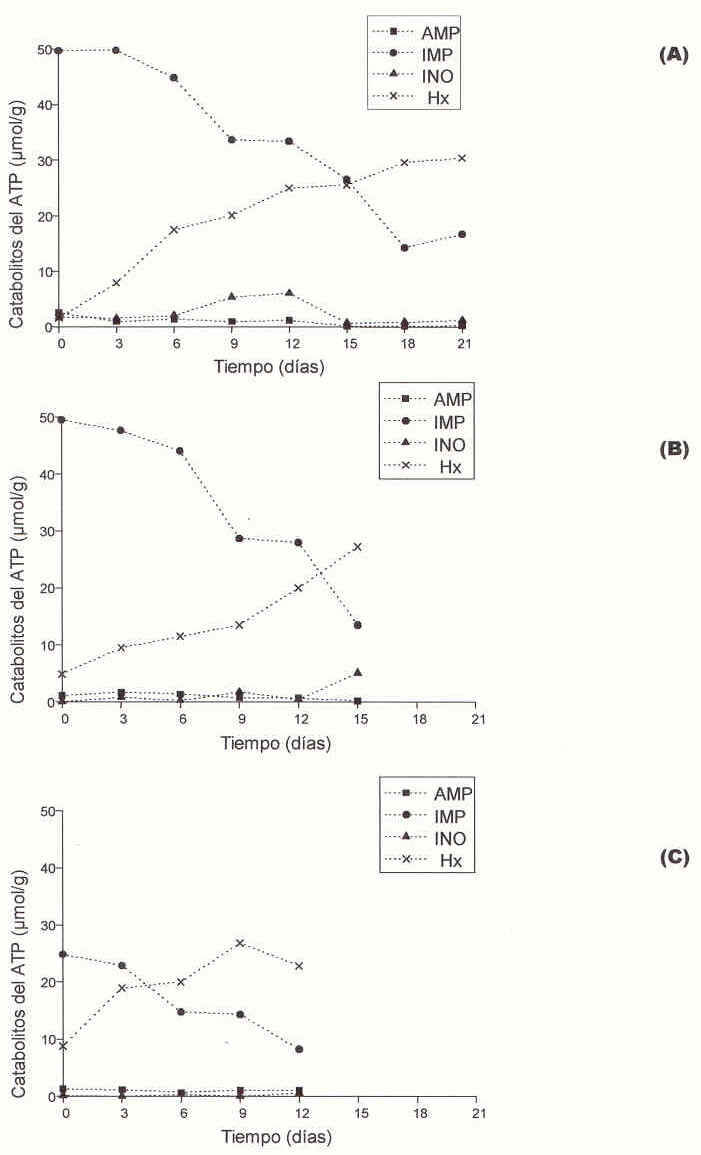
Inicialmente las concentraciones de AMP e INO fueron cercanas a 2,0.10-4 µmol/g, fluctuando muy poco a lo largo del período de almacenamiento; el comportamiento de ambos catabolitos fue independiente de la condición de TRR (TRR0h vs TRR4h, r = 0,43; TRR 0h vs TRR8h, r= 0,26; TRR4h vs TRR8h, r= -0,16, para el AMP y TRR0h vs TRR4h, r = 0,18; TRR 0h vs TRR8h, r= 0,43; TRR4h vs TRR8h, r= -0,33, para el IMP ).
Las tilapias almacenadas luego de 0h y 4h de TRR mostraron niveles iniciales de IMP significativamente mayores (p< 0,05; 5,0.10º µmol/g) que aquéllas almacenadas con 8 h de TRR (2,5.10-3 µmol/g), mientras que los niveles iniciales de Hx en éstas últimas fueron mayores (p<0,05; 9,5.10-4 µmol/g; 4,8.10-1 µmol/g; 2,3.10-4µmol/g para TRR de 8h, 4h y 0h respectivamente). Al final del almacenamiento las concentraciones de imp fueron de 16,7.10-4 µmol/g, 13,5.10-4 µmol/ g y 8,21.10-4 µmol/g para 0 h, 4 h y 8 h de TRR respectivamente; en las mismas condiciones las concentraciones de Hx fueron de 30,4.10-1 µmol/g, 27,2.10-4 µmol/g y 22,8.10—4pmol/g.

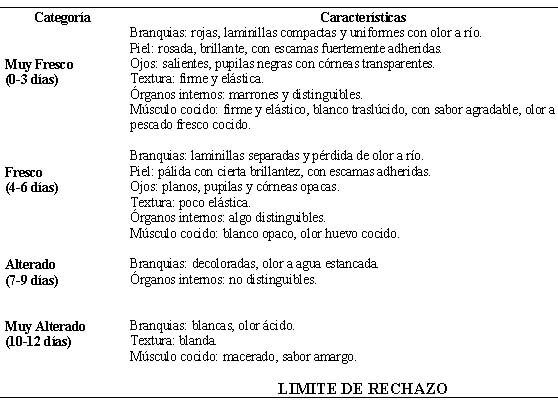
En los Cuadros 2, 3 y 4 se describen los cambios en las características sensoriales de las tilapias almacenadas en hielo a diferentes tiempos de retardo. En las tres condiciones se presentaron los mismos cambios solamente que a medida que aumentó el TRR éstos ocurrieron más tempranamente, estableciéndose los 21 días como límite de rechazo de las tilapias almacenadas con 0 h de TRR, los 15 días en las tilapias almacenadas con 4 h de TRR y los 12 días en aquéllas almacenadas con 8 h de TRR.
En términos generales al inicio del almacenamiento los ejemplares presentaron un aspecto fresco, con una piel brillante, escamas adheridas, ojos algo salientes, branquias rojas con laminillas compactas y uniformes y un olor a pescado fresco. A medida que transcurrieron los días estas características variaron, modificando el estado inicial de frescura en una y otra condición de retardo. Para los ejemplares almacenados a 00c con 0 h y 4h de retardo (Cuadros 2 y 3) prácticamente no se obtuvieron diferencias en el examen organoléptico durante los primeros 9 días de almacenamiento, posteriormente el efecto del TRR se dejó sentir en las tilapias de 4 h de retardo.
Para los ejemplares con 8 h de TRR el examen organoléptico, mostró diferencias en las características sensoriales con respecto a los otros dos tiempos de retardo, a partir del 6to día de almacenamiento (Cuadro 4).
Al relacionar la evaluación sensorial con los valores de NOB-T y PH se encontró que al 12vo día de almacenamiento en la condición de 0 h de TRR, comienzan a incrementar los valores de NBV-T (25,09 mg n%) y de pH (6,48), coincidiendo con el inicio de la pérdida de aceptabilidad organoléptica de la tilapia (r= 0,98). En la condición de 4 h de TRR este aumento se presentó el 9no día de almacenamiento (24,84 mg n%; pH = 6,47; r= 0,96) mientras que en los ejemplares almacenados con 8 h de retardo el incremento en la concentración de NBV-T (23,81 mg n%) se hizo evidente alrededor del 6to día, con un valor de pH de 6,44 y una disminución de la frescura de los ejemplares (r= 0,96).
Durante todo el período de recolección de muestras el músculo de tilapia (Oreochromis spp) se caracterizó por ser un tejido magro (<2% de grasa cruda) y con un alto porcentaje de proteínas (17,43% en promedio); adicionalmente, las variaciones de los componentes entre los lotes muestreados no fueron significativas (p>0,05) por lo que desde el punto de vista tecnológico no revisten importancia. Los lípidos de pescado son poli-insaturados lo que causa problemas de rancidez durante su almacenamiento prolongado. Adicionalmente, las proteínas miofibrilares presentes en el músculo del pescado, son las principales responsables de las propiedades emulsificantes y texturales.12

El pH muscular, en los peces vivos, es cercano a la neutralidad.9 Pedrosa-menabrito y regenstein 13 determinaron que el pH era dependiente de la especie de pescado y usualmente, inmediatamente luego del rigor mortis se encontraba entre 6,2 y 6,5. El lactato formado durante la glicólisis en el tejido muscular post-mortem disminuye el PH a partir del primer día de almacenamiento, aún a temperaturas por debajo de los 00c.14 Luego de la glicólisis, ciertos cambios autolíticos tales como el rompimiento de proteínas proporcionan las condiciones óptimas para el crecimiento y la reproducción de la microflora contaminante la cual puede producir aminas que elevan el PH del producto.15-17
En el presente estudio, el incremento del pH puede atribuirse a una serie de eventos de naturaleza enzimática y microbiológica que produjeron proteólisis y formación de compuestos alcalinos que neutralizaron el ácido láctico formado y revertieron los valores de pH final.
El incremento producido en el NBV-T indica que la condición de retardo favorece el desarrollo y crecimiento bacteriano, a partir del cual se producen los compuestos volátiles nitrogenados, lo que pone de manifiesto la importancia de colocar el pescado en hielo a la brevedad posible. Las bases volátiles totales aumentan con el incremento de la descomposición en el pescado; en peces dulce-acuícolas están representadas por el amoníaco principalmente, producido por la desaminación de los aminoácidos libres por acción bacteriana.9
En relación a los productos de degradación del ATP en el músculo dorsal de la tilapia, el AMP se determinó en niveles muy bajos en las tres condiciones de TRR estudiadas sin observarse mayores variaciones durante el período de estudio (Figura 4 ). Ha sido bien establecido que en el músculo de pescado la hidrólisis de los nucleótidos de adenina hasta el estado de IMP ocurre rápidamente. Según Okuma et al. 18 los nucleótidos de adenina desaparecen del músculo del pescado luego de 24 horas de almacenamiento. Esta desaparición es consecuencia de la desaminación y desfosforilación parcial del ATP.
Los valores iniciales de IMP disminuyeron significativamente (p<0,05) hasta el final del período de almacenamiento. El patrón de descomposición del imp depende de la especie, en muestras de sardina (Sardinella aurita), almacenada en hielo, el ATP se descompuso rápidamente a imp, alcanzando los niveles mas bajos luego de 20 días de almacenamiento19 mientras que en muestras de “Hoki” en hielo, la degradación del imp fue lenta y al final de 25 días, todavía mantenía un 39% de IMP en relación al total de nucleótidos.16
En las tres condiciones estudiadas, desde el inicio del almacenamiento ocurrió una acumulación progresiva de Hx, lo que permite sugerir que posiblemente esta especie es una formadora de este catabolito. La determinación de los niveles de Hx es utilizada para evaluar la duración del tiempo de almacenamiento en peces marINOs, debido a que su acumulación en el tejido muscular de pescado es el reflejo de la fase final de los cambios autolíticos, después de lo cual se produce un desarrollo bacteriano acelerado.20
Los niveles de INO en el músculo no incrementaron a expensas de la desaparición del IMP, ellos se mantuvieron constantes durante todo el almacenamiento. Posiblemente la falta de correspondencia entre la disminución de la concentración muscular de IMP y el aumento en la concentración de INO se debe a que la velocidad de hidrólisis de INO a Hx es mayor que la velocidad de hidrólisis de IMP a Hx.21
Los resultados obtenidos en la evaluación sensorial indicaron que el deterioro de la calidad se percibió simultáneamente en las muestras crudas que en las cocidas. Sin embargo, Manthey et al. 22 señalan que la pérdida de frescura en pescado se percibe mas tardíamente en las muestras cocidas que en las crudas debido a que con la cocción se enmascaran los cambios de calidad, especialmente los relacionados con olores desagradables pues la mayor parte de éstos son eliminados. Lo resultados obtenidos en la evaluación sensorial sugieren que la apariencia de las branquias y su olor son los mejores indicadores de frescura en las tilapias almacenadas en hielo.
Si correlacionamos los cambios en las características organolépticas con los valores de IMP e Hx se observa que el inicio de la pérdida de frescura (10-15 días en las tilapias almacenadas luego de 0 hr de TRR; 10-12 días en las tilapias almacenadas luego de 4 h de TRR; 7-9 días en las tilapias almacenadas luego de 8 h de TRR) se corresponde con una disminución en la concentración de IMP y un aumento en la concentración de Hx, lo cual es consecuencia de que la acumulación de IMP incide sobre el desarrollo de aromas y sabores agradables, mientras que la formación de Hx conlleva al desarrollo de un sabor amargo.9
Los resultados de este estudio indican que los procesos bioquímicos que influyen en el deterioro del pescado se aceleran a medida que incrementa el tiempo transcurrido entre la captura del pescado y su almacenamiento en hielo siendo la condición ideal para el almacenamiento de la tilapia (Oreochromis spp), su colocación en hielo sin demora (0h de TRR), lo que garantiza una vida útil de 21 días.
El inicio de la pérdida de la frescura de la tilapia, en las tres condiciones evaluadas, coincidió con un incremento significativo (p<0,05) en los valores de NBV-T y pH así como con una disminución del IMP y un concomitante aumento de Hx, lo que permite catalogar a Orochromis spp como formadora de este metabolito.
Los autores desean expresar su agradecimiento al consejo científico y humanístico de la Universidad Central de Venezuela (CDCH) por el financiamiento de este trabajo a través del proyecto N0 03-103840-97.